
中国全科医学 ›› 2022, Vol. 25 ›› Issue (14): 1779-1789.DOI: 10.12114/j.issn.1007-9572.2021.01.029
所属专题: 肿瘤最新文章合辑
谢馨, 陈艳, 何志旭*( ), 黄佩, 陈青华
), 黄佩, 陈青华
收稿日期:2021-06-26
修回日期:2021-09-19
出版日期:2022-03-24
发布日期:2022-04-07
通讯作者:
何志旭
基金资助:
Xin XIE, Yan CHEN, Zhixu HE*(), Pei HUANG, Qinghua CHEN
Received:2021-06-26
Revised:2021-09-19
Published:2022-03-24
Online:2022-04-07
Contact:
Zhixu HE
About author:摘要: 恶性肿瘤的发生、发展是一个不断演进的过程,其具有侵袭、浸润和转移的基本生物学特征。WASP家族作为一类新型的肌动蛋白调节蛋白,是参与维持肌动蛋白细胞骨架的基本形态的关键成分,在维持细胞基础结构、细胞运动、细胞信号传导等在内的各种细胞过程中发挥着重要作用,同时也可介导肿瘤细胞的增殖、侵袭和转移。本文综述了WASP家族在肿瘤发生、发展中的作用,以及在不同肿瘤中的表达情况及其相关信号通路,提示WASP家族在肿瘤的发生及进展中存在广阔的研究空间,对于WASP家族在各种肿瘤中的机制研究、预后预测、甚至治疗等方面均有着重要意义。
| WASP家族 | 特殊生理作用 | 共同生理作用 |
|---|---|---|
| WASP | 细胞毒性[ | 促进肌动蛋白聚合[ 调节细胞骨架、维持细胞基础结构[ 细胞迁移、运动[ 促进侵袭性伪足形成[ |
| 受体信号传导[ | ||
| 促进淋巴细胞和髓细胞迁移[ | ||
| T淋巴细胞的免疫突触形成[ | ||
| 单核细胞的吞噬[ | ||
| N-WASP | 调节肠上皮分化[ | |
| 维持正常组织结构的完整性[ | ||
| 调节成肌细胞与细胞之间的融合[ | ||
| 调节少突细胞髓鞘形成[ | ||
| 宿主-病原体相互作用[ | ||
| 寄生虫感染[ | ||
| 基质重塑过程[ | ||
| 血管内皮细胞迁移及血管新生[ | ||
| WHAMY | 维持高尔基体结构[ | |
| 维持胞内物质运输过程[ | ||
| 促进细胞自噬体形成[ | ||
| 细胞减数分裂过程[ | ||
| WASH | 细胞减数分裂过程[ | |
| YMY | 影响血睾屏障功能[ | |
| 干扰精子发生过程[ |
表1 WASP家族生理功能
Table 1 Physiological functions of WASP family
| WASP家族 | 特殊生理作用 | 共同生理作用 |
|---|---|---|
| WASP | 细胞毒性[ | 促进肌动蛋白聚合[ 调节细胞骨架、维持细胞基础结构[ 细胞迁移、运动[ 促进侵袭性伪足形成[ |
| 受体信号传导[ | ||
| 促进淋巴细胞和髓细胞迁移[ | ||
| T淋巴细胞的免疫突触形成[ | ||
| 单核细胞的吞噬[ | ||
| N-WASP | 调节肠上皮分化[ | |
| 维持正常组织结构的完整性[ | ||
| 调节成肌细胞与细胞之间的融合[ | ||
| 调节少突细胞髓鞘形成[ | ||
| 宿主-病原体相互作用[ | ||
| 寄生虫感染[ | ||
| 基质重塑过程[ | ||
| 血管内皮细胞迁移及血管新生[ | ||
| WHAMY | 维持高尔基体结构[ | |
| 维持胞内物质运输过程[ | ||
| 促进细胞自噬体形成[ | ||
| 细胞减数分裂过程[ | ||
| WASH | 细胞减数分裂过程[ | |
| YMY | 影响血睾屏障功能[ | |
| 干扰精子发生过程[ |
| 作用 | 机制 |
|---|---|
| 参与细胞黏附 | 激活Arp2/3复合物,促进肌动蛋白结构形成[ |
| WasC通过阻断VCA结构域的活性抑制F-肌动蛋白聚合从而抑制细胞之间的黏附作用[ | |
| 降解细胞外基质 | WAVE3激活NFκB通路和MAPK信号通路,从而激活基质金属蛋白酶MMP[ |
| 促进血管新生 | WASP重组细胞骨架,促进血管内皮细胞移行[ |
| WASP通过激活MMP,启动内皮细胞的激活和迁移,稳定新生血管[ | |
| N-WASP磷酸化从而促进血管内皮细胞迁移及血管新生[ | |
| 促进细胞运动 | N-WASP在甲状腺激素刺激下,激活FAK/paxillin/cortactin/N-WASP/Arp2/3复合信号通路,增强细胞运动能力[ |
| 肌动球蛋白介导的细胞收缩为细胞体向前运动提供能量,为胞体前缘产生收缩力,从而促进胞体与基质分离[ | |
| 促进形成侵袭性伪足 | 在Rac和Cdc42调节下,促进质膜突起形成层状伪足和丝状伪足[ |
| 参与肿瘤免疫 | 参与免疫突触的形成和T淋巴细胞发育[ |
| 调节BCR信号通路和抗原内化提呈过程[ | |
| 影响巨噬细胞的趋化作用和吞噬作用[ | |
| N-WASP通过NF-κB和MAPK的信号通路,影响炎性因子表达水平[ | |
| 参与肿瘤代谢 | 与TRIM21形成螯合物,从而保持PFK的活性,促进糖酵解过程[ |
| 通过Rac1/ROS信号通路促进活性氧(ROS)的生成[ | |
| 参与肿瘤死亡 | WAVE3激活AKT促存活信号通路,加强肿瘤细胞对凋亡信号和TNF-α介导的细胞死亡信号的敏感性,从而促进肿瘤细胞的凋亡和死亡[ |
| WAVE2通过Rac1/ROS信号通路促进活性氧(ROS)生成[ | |
| WASP缺陷时,在CD24交联诱导下使细胞凋亡增强[ | |
| 促进自噬小体形成[ | |
| 参与肿瘤耐药 | 通过Beclin1/Bcl-2和Beclin1/PI3K-Ⅲ复合物途径增强白血病细胞的存活和调节化疗耐药[ |
表2 WASP家族在肿瘤中的作用及机制
Table 2 Role and mechanism of WASP family in cancer formation and progression
| 作用 | 机制 |
|---|---|
| 参与细胞黏附 | 激活Arp2/3复合物,促进肌动蛋白结构形成[ |
| WasC通过阻断VCA结构域的活性抑制F-肌动蛋白聚合从而抑制细胞之间的黏附作用[ | |
| 降解细胞外基质 | WAVE3激活NFκB通路和MAPK信号通路,从而激活基质金属蛋白酶MMP[ |
| 促进血管新生 | WASP重组细胞骨架,促进血管内皮细胞移行[ |
| WASP通过激活MMP,启动内皮细胞的激活和迁移,稳定新生血管[ | |
| N-WASP磷酸化从而促进血管内皮细胞迁移及血管新生[ | |
| 促进细胞运动 | N-WASP在甲状腺激素刺激下,激活FAK/paxillin/cortactin/N-WASP/Arp2/3复合信号通路,增强细胞运动能力[ |
| 肌动球蛋白介导的细胞收缩为细胞体向前运动提供能量,为胞体前缘产生收缩力,从而促进胞体与基质分离[ | |
| 促进形成侵袭性伪足 | 在Rac和Cdc42调节下,促进质膜突起形成层状伪足和丝状伪足[ |
| 参与肿瘤免疫 | 参与免疫突触的形成和T淋巴细胞发育[ |
| 调节BCR信号通路和抗原内化提呈过程[ | |
| 影响巨噬细胞的趋化作用和吞噬作用[ | |
| N-WASP通过NF-κB和MAPK的信号通路,影响炎性因子表达水平[ | |
| 参与肿瘤代谢 | 与TRIM21形成螯合物,从而保持PFK的活性,促进糖酵解过程[ |
| 通过Rac1/ROS信号通路促进活性氧(ROS)的生成[ | |
| 参与肿瘤死亡 | WAVE3激活AKT促存活信号通路,加强肿瘤细胞对凋亡信号和TNF-α介导的细胞死亡信号的敏感性,从而促进肿瘤细胞的凋亡和死亡[ |
| WAVE2通过Rac1/ROS信号通路促进活性氧(ROS)生成[ | |
| WASP缺陷时,在CD24交联诱导下使细胞凋亡增强[ | |
| 促进自噬小体形成[ | |
| 参与肿瘤耐药 | 通过Beclin1/Bcl-2和Beclin1/PI3K-Ⅲ复合物途径增强白血病细胞的存活和调节化疗耐药[ |
| 肿瘤 | WASP家族 | 表达 | 机制 | 预后作用 | 通路 | |
|---|---|---|---|---|---|---|
| 血液系统 | CML | WASP | 随CML的进展而降低,与BCR-ABL1的表达呈负相关[ | 不明 | 不良预后[ | 不明 |
| JMML | WAS基因突变 | 下降[ | 不明 | 不详 | 可能与RAS通路活化相关[ | |
| 淋巴瘤 | WASP选择性下降[ | 下降[ | WASP蛋白缺陷时,CDC42与GTP结合增加,激活下游STAT3和MAPK通路,加速T淋巴瘤的病程[ | 不良预后[ | STAT3通路、换行MAPK通路[ | |
| 消化系统 | 肝细胞癌 | N-WASP | 升高[ | 作为细胞骨架调节因子,促进肿瘤细胞侵袭和转移[ | 不良预后[ | 不明 |
| WAVE2 | 升高[ | 作为细胞骨架调节因子,促进肿瘤细胞侵袭和转移[ | 不良预后[ | 不明 | ||
| 肝内胆管细胞癌 | WAVE3 | 升高[ | 诱导上皮-间充质转化[ | 不良预后[ | 不明 | |
| 胰腺导管细胞癌 | WAVE3 | 升高[ | 抑制PDK2的表达,抑制AKT通路中Ser473的磷酸化,从而影响蛋白激酶B(PBK/AKT)通路,影响AKT途径下游蛋白如EMT相关蛋白、p53、Bcl-2和cyclin D1的表达,影响细胞增殖、迁移和侵袭[ | 不良预后[ | AKT途径[ | |
| N-WASP | 升高[ | 通过p120-catenin/β-catenin/p21通路,在胰腺导管细胞癌中发挥致癌作用[ | 不良预后[ | p120-catenin/β-catenin/p21通路[ | ||
| 大肠癌 | WASP | 低表达甚至无表达[ | 不详 | 不详 | 不详 | |
| N-WASP | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| 食管癌 | WAVE3 | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | |
| WASH | 升高[ | WASH在miR-637作用下表达上调,进而上调CXCL8,促进食管鳞癌细胞的干性,促进癌细胞的生长及转移[ | 不良预后[ | 不详 | ||
| N-WASP | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| 生殖系统 | 乳腺癌 | WASP | 下降[ | WASP的缺陷会影响集落刺激因子对巨噬细胞的趋化作用,从而打断巨噬细胞与乳腺癌细胞之间的作用,使肿瘤细胞恶性增殖,促进癌细胞的运动、侵袭、内渗及转移[ | 不良预后[ | 不详 |
| N-WASP | 下降[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| WAVE2[ | 升高[ | 通过Rac1-WAVE信号通路促进侵袭性板足形成,从而促进肿瘤细胞的侵袭和迁移能力[ | 不良预后[ | Rac1-WAVE信号通路[ | ||
| 卵巢癌 | WAVE | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | |
| 前列腺癌 | WAVE1 | 升高[ | 肌动蛋白骨架机制,促进运动、侵袭和转移[ | 不良预后[ | 不详 | |
| WAVE3 | 升高[ | 肌动蛋白骨架机制,促进运动、侵袭和转移[ | 不良预后[ | 不详 | ||
| WASP | 升高[ | WASP可接收Rho-GTPase家族的上游信号,将信号传递给Arp2/3复合体,从而导致细胞前缘肌动蛋白的快速聚合,促进侵袭性伪足的形成从而促进癌细胞转移[ | 不良预后[ | 不详 | ||
| 宫颈癌 | N-WASP[ | 升高[ | N-WASP通过调节p38 MAPK通路的活性,参与肿瘤浸润与转移[ | 不良预后[ | p38-MAPK通路[ | |
| 子宫内膜癌 | N-WASP | 升高[ | 在miR-216a-5p调控下表达上调,从而促进子宫内膜癌细胞的增殖、迁移和侵袭能力[ | 不良预后[ | miR-216a-5p调控[ | |
| 泌尿系统 | 肾癌 | N-WASP | 下降[ | SNHG14通过与转录因子SP1结合使其在癌组织中表达增强,并通过miR-203依赖的方式调节N-WASP蛋白表达水平,从而促进肾脏肿瘤细胞迁移和侵袭的能力[ | 表达升高时预示不良预后[ | 不详 |
| 呼吸系统 | 肺癌 | N-WASP | 升高[ | N-WASP可与生长阻滞特异蛋白7(GAS7)结合,从而激活纤维连接蛋白/整合素/FAK通路(N-WASP/FAK/F-actin),促进细胞骨架动力学,增加癌细胞运动,促进肺癌转移[ | 不良预后 | 纤维连接蛋白/整合素/FAK通路(N-WASP/FAK/F-actin)[ |
| 鼻咽癌 | N-WASP | 升高[ | 在CDC42结合蛋白(CIP4)激活下,促进侵袭性伪足的形成,并激活EGFR信号,从而诱导下游MMP2(基质金属蛋白酶2)上调,促进鼻咽癌转移[ | 不良预后[ | EGFR信号[ | |
| 中枢神经系统 | 神经胶质母细胞瘤 | N-WASP | 升高[ | N-WASP可参与室管膜细胞和神经元的迁移和突触调节[ | 不良预后[ | 不详 |
| 其他系统 | 黑色素瘤 | WAVE1\WAVE2 | 升高[ | WAVE1和WAVE2作为Rac下游的主要效应因子,通过激活Arp2/3复合物调节肌动蛋白细胞骨架,促进肿瘤细胞侵袭和转移[ | 不良预后[ | Rac1-WAVE信号通路[ |
表3 WASP家族在各种肿瘤中的表达及预后作用和相关通路
Table 3 Expression,prognostic role and related pathways of WASP family in various cancers
| 肿瘤 | WASP家族 | 表达 | 机制 | 预后作用 | 通路 | |
|---|---|---|---|---|---|---|
| 血液系统 | CML | WASP | 随CML的进展而降低,与BCR-ABL1的表达呈负相关[ | 不明 | 不良预后[ | 不明 |
| JMML | WAS基因突变 | 下降[ | 不明 | 不详 | 可能与RAS通路活化相关[ | |
| 淋巴瘤 | WASP选择性下降[ | 下降[ | WASP蛋白缺陷时,CDC42与GTP结合增加,激活下游STAT3和MAPK通路,加速T淋巴瘤的病程[ | 不良预后[ | STAT3通路、换行MAPK通路[ | |
| 消化系统 | 肝细胞癌 | N-WASP | 升高[ | 作为细胞骨架调节因子,促进肿瘤细胞侵袭和转移[ | 不良预后[ | 不明 |
| WAVE2 | 升高[ | 作为细胞骨架调节因子,促进肿瘤细胞侵袭和转移[ | 不良预后[ | 不明 | ||
| 肝内胆管细胞癌 | WAVE3 | 升高[ | 诱导上皮-间充质转化[ | 不良预后[ | 不明 | |
| 胰腺导管细胞癌 | WAVE3 | 升高[ | 抑制PDK2的表达,抑制AKT通路中Ser473的磷酸化,从而影响蛋白激酶B(PBK/AKT)通路,影响AKT途径下游蛋白如EMT相关蛋白、p53、Bcl-2和cyclin D1的表达,影响细胞增殖、迁移和侵袭[ | 不良预后[ | AKT途径[ | |
| N-WASP | 升高[ | 通过p120-catenin/β-catenin/p21通路,在胰腺导管细胞癌中发挥致癌作用[ | 不良预后[ | p120-catenin/β-catenin/p21通路[ | ||
| 大肠癌 | WASP | 低表达甚至无表达[ | 不详 | 不详 | 不详 | |
| N-WASP | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| 食管癌 | WAVE3 | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | |
| WASH | 升高[ | WASH在miR-637作用下表达上调,进而上调CXCL8,促进食管鳞癌细胞的干性,促进癌细胞的生长及转移[ | 不良预后[ | 不详 | ||
| N-WASP | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| 生殖系统 | 乳腺癌 | WASP | 下降[ | WASP的缺陷会影响集落刺激因子对巨噬细胞的趋化作用,从而打断巨噬细胞与乳腺癌细胞之间的作用,使肿瘤细胞恶性增殖,促进癌细胞的运动、侵袭、内渗及转移[ | 不良预后[ | 不详 |
| N-WASP | 下降[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | ||
| WAVE2[ | 升高[ | 通过Rac1-WAVE信号通路促进侵袭性板足形成,从而促进肿瘤细胞的侵袭和迁移能力[ | 不良预后[ | Rac1-WAVE信号通路[ | ||
| 卵巢癌 | WAVE | 升高[ | 肌动蛋白骨架机制[ | 不良预后[ | 不详 | |
| 前列腺癌 | WAVE1 | 升高[ | 肌动蛋白骨架机制,促进运动、侵袭和转移[ | 不良预后[ | 不详 | |
| WAVE3 | 升高[ | 肌动蛋白骨架机制,促进运动、侵袭和转移[ | 不良预后[ | 不详 | ||
| WASP | 升高[ | WASP可接收Rho-GTPase家族的上游信号,将信号传递给Arp2/3复合体,从而导致细胞前缘肌动蛋白的快速聚合,促进侵袭性伪足的形成从而促进癌细胞转移[ | 不良预后[ | 不详 | ||
| 宫颈癌 | N-WASP[ | 升高[ | N-WASP通过调节p38 MAPK通路的活性,参与肿瘤浸润与转移[ | 不良预后[ | p38-MAPK通路[ | |
| 子宫内膜癌 | N-WASP | 升高[ | 在miR-216a-5p调控下表达上调,从而促进子宫内膜癌细胞的增殖、迁移和侵袭能力[ | 不良预后[ | miR-216a-5p调控[ | |
| 泌尿系统 | 肾癌 | N-WASP | 下降[ | SNHG14通过与转录因子SP1结合使其在癌组织中表达增强,并通过miR-203依赖的方式调节N-WASP蛋白表达水平,从而促进肾脏肿瘤细胞迁移和侵袭的能力[ | 表达升高时预示不良预后[ | 不详 |
| 呼吸系统 | 肺癌 | N-WASP | 升高[ | N-WASP可与生长阻滞特异蛋白7(GAS7)结合,从而激活纤维连接蛋白/整合素/FAK通路(N-WASP/FAK/F-actin),促进细胞骨架动力学,增加癌细胞运动,促进肺癌转移[ | 不良预后 | 纤维连接蛋白/整合素/FAK通路(N-WASP/FAK/F-actin)[ |
| 鼻咽癌 | N-WASP | 升高[ | 在CDC42结合蛋白(CIP4)激活下,促进侵袭性伪足的形成,并激活EGFR信号,从而诱导下游MMP2(基质金属蛋白酶2)上调,促进鼻咽癌转移[ | 不良预后[ | EGFR信号[ | |
| 中枢神经系统 | 神经胶质母细胞瘤 | N-WASP | 升高[ | N-WASP可参与室管膜细胞和神经元的迁移和突触调节[ | 不良预后[ | 不详 |
| 其他系统 | 黑色素瘤 | WAVE1\WAVE2 | 升高[ | WAVE1和WAVE2作为Rac下游的主要效应因子,通过激活Arp2/3复合物调节肌动蛋白细胞骨架,促进肿瘤细胞侵袭和转移[ | 不良预后[ | Rac1-WAVE信号通路[ |
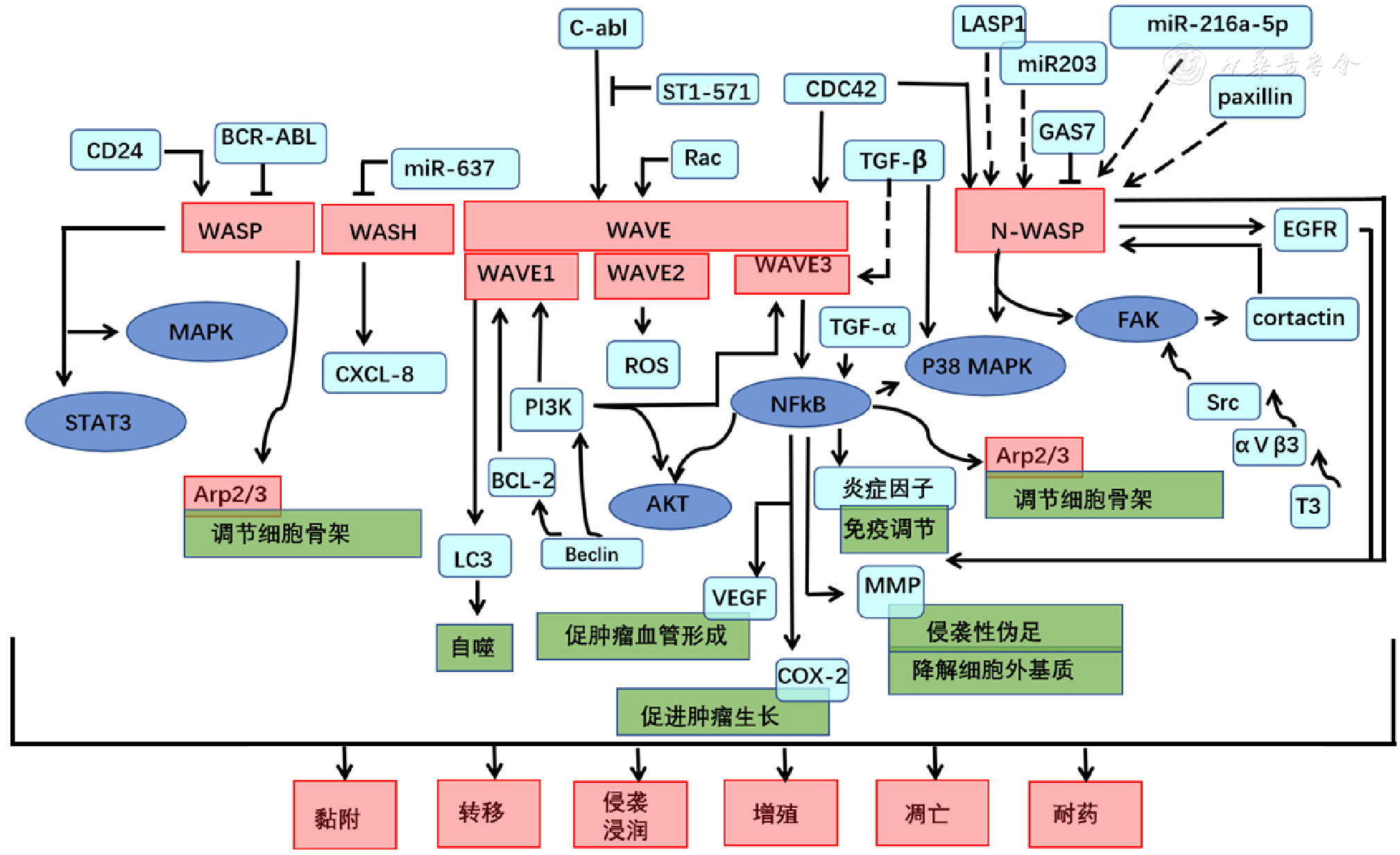
图1 WASP相关信号通路示意图注:PI3K=磷脂酰肌醇3激酶,TGF=转化生长因子,Arp=肌动蛋白调节因子,CXCL=趋化因子
Figure 1 Schematic diagram of signaling pathways mediated by WASP familyproteins in cancer formation and progression
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
王蓓蓓,张丽莙. IQGAP1信号通路在肺炎衣原体感染促进血管新生中的调控作用[J]. 中国病理生理杂志,2015,31(10):1814,1815. DOI:10.3969/j.issn.1000-4718.2015.10.100.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
范嘉盈. JMY蛋白在Sertoli细胞中的功能及其作用机制研究[D]. 上海:上海交通大学,2018.
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
申丽媛,唐良萏,张婧,等. WAVEs亚家族在卵巢癌中的表达及临床意义[J]. 基础医学与临床,2013,33(9):1135-1140. DOI:10.16352/j.issn.1001-6325.2013.09.022.
|
| [76] |
秦巧红,张楠,赵书君,等. miR-216a-5p和WASL在子宫内膜癌组织中的表达及其调控子宫内膜癌细胞增殖、迁移和侵袭的分子机制[J]. 吉林大学学报:医学版,2020,46(4):844-850. DOI:10.13481/j.1671-587x.20200429.
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
鲁萍,董吉顺,平金良,等. N-WASP在肾透明细胞癌组织及癌旁正常肾组织中的表达及与肿瘤病理分级和临床分期的相关性分析[J]. 中华医院感染学杂志,2017,27(20):4668-4671. DOI:10.11816/cn.ni.2017-170834.
|
| [82] |
|
| [83] |
陈健,纪志刚,刘广华,等. 肾透明细胞癌N-WASP蛋白表达特点及与临床病理相关性[J]. 基础医学与临床,2015,35(3):366-370. DOI:10.16352/j.issn.1001-6325.2015.03.016.
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [1] | 冯宇茜, 王津津, 蔡一, 朱润芝, 朱勤. 乳酸化修饰在肾脏疾病中的研究进展[J]. 中国全科医学, 2026, 29(21): 3056-3063. |
| [2] | 刘帅, 苑世朝, 刘琰, 贾硕贤, 赵岩, 王紫怡, 周春华, 于静. 两性霉素B治疗侵袭性真菌病的疗效、安全性及经济性评价:一项回顾性队列研究[J]. 中国全科医学, 2026, 29(21): 2967-2974. |
| [3] | 李文萍, 陈建华, 许家培, 金雪, 潘子涵, 迟春花. 数智化健康管理平台赋能社区老年人健康服务的范围综述[J]. 中国全科医学, 2026, 29(21): 2938-2949. |
| [4] | 程玉, 韩士尧, 孙平, 向娟, 杨辉, 李东泽, 郑莉, 廖晓阳. 慢性病管理中的社会经济与人口学挑战分析[J]. 中国全科医学, 2026, 29(20): 2800-2807. |
| [5] | 陈义萍, 张剑梅, 厉彦虎, 李睿, 张瑞平, 杨萍. COVID-19后遗症心肌损伤的诊疗进展[J]. 中国全科医学, 2026, 29(18): 2585-2592. |
| [6] | 张明悦, 侯艳. 呼出气、肺泡气一氧化氮在间质性肺疾病中的研究进展[J]. 中国全科医学, 2026, 29(18): 2571-2576. |
| [7] | 马晓雨, 马晓梅, 武国智, 郑亚, 郭庆红. 中性粒细胞外陷阱在胃癌中的研究进展[J]. 中国全科医学, 2026, 29(17): 2425-2432. |
| [8] | 谢丹翎, 刘博雅, 李晓光, 王瀚苇, 马强, 方靖琴, 王舒楠. 基于超声生境成像对乳腺叶状肿瘤良恶性鉴别的效能研究[J]. 中国全科医学, 2026, 29(17): 2368-2375. |
| [9] | 李佳威, 葛奥淇, 高欣怡, 李娟娟, 袁蓓蓓. 医防融合的概念内涵和实现路径:一项系统综述研究[J]. 中国全科医学, 2026, 29(16): 2156-2166. |
| [10] | 汪洋, 金花, 杨森, 杨辉, 于德华. 中国基本医疗服务功能特征及其对卫生服务结局的影响路径:一项15年的混合方法系统综述[J]. 中国全科医学, 2026, 29(16): 2140-2155. |
| [11] | 刘婧琳, 李维, 张晓龙, 宋晓坤, 周西蓓, 王冠元, 张洁. 医药联合在弥漫大B细胞淋巴瘤合并急性肾功能衰竭患者中的治疗实践与分析[J]. 中国全科医学, 2026, 29(12): 1599-1606. |
| [12] | 叶鑫瑛, 殷安康, 刘定岭, 吕翔, 王逸. 按病种分值付费支付模式下综合医院全科医学科发展研究[J]. 中国全科医学, 2026, 29(11): 1378-1384. |
| [13] | 施雨峰, 卢晨霞, 吕安淇, 李晓东. 基于肠-肝轴理论探讨韦荣球菌属与肝脏疾病关系的研究进展[J]. 中国全科医学, 2026, 29(11): 1488-1496. |
| [14] | 孙宇辰, 石雅馨, 师伟. 经血源性干细胞联合补肾活血类中药治疗子宫内膜功能减退类疾病的研究进展[J]. 中国全科医学, 2026, 29(11): 1481-1487. |
| [15] | 王昱棋, 叶睿雪, 高焱, 薛凯文, 周静, 李冬霞, 郝英姿, 李晓轩, 王玉龙. 康复分级诊疗体系的国内外发展现状及应用需求探讨[J]. 中国全科医学, 2026, 29(10): 1250-1255. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||










